
Remember me

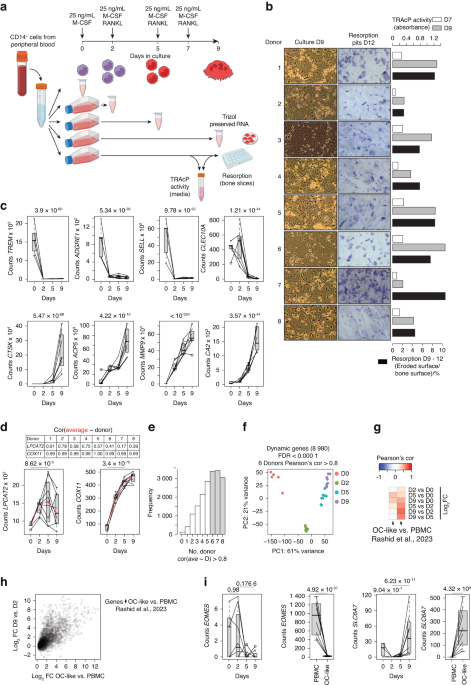
To investigate the functions of α-parvin in long bone development, we generated PARVAflox/floxPrx1cre mice (cKO mice), in which α-parvin was ablated from the limb bud mesoderm. The cKO mice, irrespective of male or female, died before two months of age. In addition, cKO mice were produced at a ratio lower than that Mendelian law predicted, possibly due to localization of the genes encoding α-parvin and Prx1-Cre in the same chromosome, as both genes are known to localize in chromosome 7.28
The cKO mice showed significant dwarfism starting from the adolescent stage (Fig. 1a). They were 15% shorter in body length at seven weeks of age compared to control mice (Fig. 1b). As for the long bone development, the tibia of cKO mice was similar in length to control at E16.5 (Fig. 1c, g) but became ~10% shorter than that of control mice at P0 (postnatal day 0) (Fig. 1d, h). The difference in tibia length became more evident in elder cKO and control mice. P7 and P30 cKO mice all showed a 15% to 20% reduction in tibia length compared to control mice (Fig. 1e, f, i, j). These results suggest that α-parvin is required for proper long bone growth, especially at the postnatal stage.
Fig. 1
Loss of α-Parvin leads to dwarfism and shorter long bones in mice. a Gross morphology of six weeks-old control and PARVAflox/floxPrx1cre mice (cKO mice). Scale bar = 1 cm. b Comparison of body length between control and cKO mice at 7 weeks of age. **P < 0.01. paired t-test. mean ± s.e.m. n = 5 mice for each group. c–f Stereoscopic images of tibia from E16.5, P0, P7, and P30 control (left) and cKO mice (right). Scale bars = 500 μm, 500 μm, 1 mm, 2 mm respectively (from left to right). g Comparison of tibia length between control and cKO embryos at E16.5. ns not significant. unpaired t-test. mean ± s.e.m. n = 3 control and 8 cKO embryos from the same litter. h Comparison of tibia length between control and cKO mice at P0. ****P < 0.000 1. paired t-test. mean ± s.e.m. n = 6 mice for each group. i Comparison of tibia length between control and cKO mice at P7. ****P < 0.000 1. paired t-test. mean ± s.e.m. n = 6 mice for each group. j Comparison of tibia length between control and cKO mice at P30. **P < 0.01. paired t-test. mean ± s.e.m. n = 3 mice for each group
Limb bud deletion of α-parvin causes widening of long bonesWe next analyzed the width of the long bones and the growth plate cartilage of the cKO mice. In the tibia, measured at the chondro-osseous junction, we found a 15% increase in width compared to control at P0 (Fig. 2a, b). Interestingly, the fibula, a thin long bone that runs aside the tibia, behaved pronounced widening in cKO mice compared to control (Fig. 2c–h). Even at E16.5, when the difference in length was not yet evident, the fibula of cKO pups was already 15% wider than control (Fig. 2c, f). The difference in width further increased to 50% at P0 (Fig. 2d, g). The widening phenotypes diminished at P7, but a 30% increase could still be observed in the fibula of cKO pups compared to control (Fig. 2e, h). These data suggest that loss of α-parvin leads to excessive horizontal growth of long bones at the embryonic and neonatal stages.
Fig. 2
Loss of α-Parvin causes widening of long bones. a Stereoscopic image of knee joints of control and cKO mice at P0. Scale bar = 1 mm. b Comparison of the tibia width between control and cKO group, *P < 0.05. paired t-test. mean ± s.e.m. n = 4 mice for each group (c–e) Stereoscopic images of tibia from E16.5, P0, P7, and P30 control (left) and cKO mice (right). Scale bars = 500 μm, 500 μm, 1 mm respectively (from left to right). Double headed arrows denote the width of fibula. f Comparison of fibula width between control and cKO embryos at E16.5. **P < 0.01. unpaired t-test. mean ± s.e.m. n = 3 control and 8 cKO embryos from the same litter. g Comparison of fibula width between control and cKO mice at P0. ****P < 0.000 1. paired t-test. mean ± s.e.m. n = 8 mice for each group. h Comparison of fibula width between control and cKO mice at P7. **P < 0.01. paired t-test. mean ± s.e.m. n = 5 mice for each group
Loss of α-parvin causes disorganization of the growth platesAnalyses of the growth plates of cKO mice revealed strikingly disorganized columnar structures. At P0 and P7, chondrocytes in the proliferative zone were normally arranged into columns that consist of at least 5 cells in control growth plates (Fig. 3a, b). In contrast, multiple short columns with less than five cells were found in the proliferative zone of cKO growth plates (Fig. 3a, b). At P30, similar columnar disorganization was found in the proliferative zone of cKO growth plates (Fig. 3a, b). Unexpectedly, the resting zone of cKO growth plates became dilated, indicating possible expansion of the stem cell population (Fig. 3a, b). In all stages, chondrocytes in the proliferative zone of cKO growth plates were much less elongated than control, as shown by the smaller aspect ratio (Fig. 3c). However, the orientation of chondrocytes only slightly increased in the cKO group (Fig. 3d). Moreover, the proliferative zone of cKO growth plates exhibited increased aggregation of chondrocytes. Around 5% of chondrocytes were found within an aggregate (Fig. S1A, B). This is confirmed by immunostaining with antibodies for pan-cadherin, which showed significant increase of cell-cell junctions in the proliferative zones of the cKO growth plates (Fig. S1C, D). These results suggest that loss of α-parvin causes disorganization of the growth plate cartilage.
Fig. 3
Loss of α-Parvin causes disorganization of growth plates. a H&E staining of the proximal tibia growth plate sections from control and cKO mice at P0, P7, and P30. Scale bars = 100 μm. b Staining of actin cytoskeleton. Sections of the proximal tibia growth plate were stained with Phalloidin (green) and counterstained with DAPI (blue). Scale bars = 20 μm. Quantification of the aspect ratio (c) and orientation (d) of chondrocytes in the proliferative zones of control and cKO growth plates at P0, P7, and P30. ns, not significant. **P < 0.01. ****P < 0.000 1. unpaired t-test. mean ± s.e.m. n > 40 cells for each group
α-parvin regulates chondrocyte rotationLive imaging of the growth plates is extremely valuable for monitoring chondrocytes’ behavior during long bone development. In chicks, the ex-vivo culture of the metatarsal or metacarpal has been used to study the mechanism of long bone growth and gene functions.5,29,30 In mice, Sarah et al. first applied live imaging on newborn pre-sphenoidal synchondrosis, a piece of tiny and flat cartilage in the cranial base, using Rosa26mTmG(mTmG) transgenic mice.31 Keisho Hirota et al. also applied two-photon live imaging in culturing embryonic murine ulna to track the growth of long bones.32 To elucidate the cellular mechanism underlying the broader but shorter long bones of cKO mice, we employed two-photon live imaging and a neonatal fibula explant culture model to analyze the behavior of chondrocytes in situ. The fibula is a thin long bone that runs aside the tibia, with relatively flat growth plates. The surrounding tissues of the growth plates are thin, allowing two-photon microscopy of the chondrocytes. By combining two-photon live imaging and fibula explant culture, we could visualize chondrocyte rotation events within neonatal growth plates. In both control mice bearing mTmG reporters (mTmG-control mice) and Rosa26mTmG/+; Prx1-Cre, Parvaflox/flox mice (mTmG-cKO mice), consistent with the previous study,31 we found that the chondrocytes in the proliferative zone of fibula growth plates underwent a rotational movement process after cell division (Movies S1–S4). All daughter cells formed cell-cell adhesion after cytokinesis in both mTmG-control and mTmG-cKO group (Movies S1–S18). The daughter chondrocytes then pivoted along the cell adhesion surface and adjusted to the proper orientation, which could be parallel, orthogonal, or at a certain angle to the x-axis (chondro-osseous junction) (Fig. 4a, b, Movies S1–S18). We counted more than 300 events from at least three independent experiments for each group to quantify the chondrocyte rotation.
Fig. 4
Live imaging of the growth plates in a fibula explant culture. a, b Snapshots from live imaging of chondrocytes undergoing chondrocyte rotation in mTmG-control or mTmG-cKO proximal fibula growth plates. The red rectangles highlight the dividing and rotating chondrocytes. NR normal rotation, IR incomplete rotation, FR failed rotation. GFP signals were excited with 945 nm laser. The boxes at the lower right of each image indicate the elapsed time. The percentages of each type of rotation events are shown on the right. The number of the corresponding type out of the total number of rotation events was shown in the brackets. In total, 371 rotation events of explant cultures from five mTmG-control mice and 382 rotation events of explant cultures from four cKO mice were counted. Scale bar = 20 μm
In mTmG-control growth plates, most (81%) of chondrocytes normally rotated (NR), with a final orientation of cell-cell adhesion parallel or at an angle less than 30 degrees to the x-axis (Figs. 4a and 5a, b, Movies S1–S4). 10% of chondrocyte rotation events were incomplete (IR) in mTmG-control as the final angles of the cell-cell adhesion to the x-axis were more than 30 degrees (but less than 60 degrees) (Figs. 4a and 5a, b, Movie S5). 9% of mTmG-control chondrocytes did not rotate after cell division or stopped rotation at an angle greater than 60 degrees. These cells were classified as "Failed rotation" (FR) (Figs. 4a and 5a, b, Movie S6).
Fig. 5
α-Parvin regulates chondrocyte rotation. a Classification of chondrocyte rotation events based on the final angles of cell-cell adhesion. Theoretically, the cell-cell adhesion bisects the major axis (green dashed line) after cytokinesis. Red dashed lines indicate the horizontal axis (x-axis) of long bones. b Summary of the outcomes of chondrocyte rotation events in mTmG-control and mTmG-cKO group. n = 5 explant cultures for mTmG-control group, n = 4 explant cultures for mTmG-cKO group. ****P < 0.000 1, unpaired t-test, mean ± s.e.m. Tracing of the angles of cell-cell adhesions during chondrocyte rotation in mTmG-control (c) and mTmG-cKO (d) group for up to 10 h. n = 25 rotation events for each group. e, f Angles of cell-cell adhesions at the beginning and at the end of the chondrocyte rotations. n = 25 rotation events for each group
In contrast, in mTmG-cKO mice, a much larger percentage (48%) of growth plate chondrocytes failed to rotate (Figs. 4b and 5a, b, Movies S13–S16). Furthermore, incomplete rotation was observed in 29% of mTmG-cKO chondrocytes (Figs. 4b and 5a, b, Movies S9–S12). Only 23% of mTmG-cKO chondrocytes underwent normal rotation after cell division (Figs. 4b and 5a, b, Movies S7 and S8). By tracing the angles of cell-cell adhesion during the rotation process, we found that most chondrocytes in mTmG-control growth plates completed rotation within four hours, and the angles of cell-cell adhesion quickly converged to around 0° to the x-axis (Fig. 5c, e). Concomitantly, the daughter chondrocytes became flattened (Fig. 5c and Fig. S3A). By contrast, the angles of the cell-cell junction in most of the mTmG-cKO chondrocytes barely changed even after 10 hours (Fig. 5d, f). Since most mTmG-cKO chondrocytes did not undergo significant rotation, the flattening of the cells was also impaired (Fig. 5d and Fig. S3b). The net change of cell-cell adhesion angles also dramatically decreased to one-third of that in mTmG-control group (Fig. S3C).
Of note, 7% of mTmG-cKO chondrocytes were found to form aggregates during the live imaging experiments (Fig. S2A). One of the chondrocytes in a doublet divided before it separated from the sister cell. The doublet thus became a triplet and had three intercellular junction interfaces. They eventually adapted a three-pointed star style (Fig. S2A). Therefore, all three daughter cells failed to stack into the column. Indeed, mTmG-cKO growth plates contained chondrocyte aggregates within the proliferative zone, indicating its possible contribution to disorganized columns and wider long bones (Fig. S1A–D). By contrast, no aggregate formation was observed in mTmG-control chondrocytes. Taken together, with two-photon time-lapse imaging, our results suggest that α-parvin regulates chondrocyte rotation, which is necessary for the formation of the columnar structure.
Loss of α-parvin induces multiple chondrocyte defectsIn growth plate cartilage, apoptosis typically occurs within the hypertrophic zone and borderline area.33 To assess the effect of α-parvin deficiency on cell survival, we performed Terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) staining on control and cKO growth plates. Although no change of apoptosis was found between the control and cKO hypertrophic zone and the lower borderline area, we detected significant increase of apoptosis in the cKO growth plates compared to that of the control at P7 (Fig. 6a). Most TUNEL signals were detected in the lateral region of cKO resting zone (Fig. 6a). In addition to elevated cell death, we also found increased binucleation at the late neonatal stage (P5-P7) in cKO growth plates. The percentage of binucleated cKO chondrocytes was similar to that of the control at P0 but it was increased significantly (to 8%) at P5-P7 (Fig. S4E, F). BrdU incorporation assays showed that the percentage of BrdU-positive nuclei of cKO proliferative zones were comparable with that of control, indicating that the entry into S phase was unaffected in cKO chondrocytes at the neonatal and embryonic stages (Fig. S4A–D).
Fig. 6
Loss of α-parvin results in increased apoptosis and dilated resting zone. a Apoptosis. Sections of the P7 proximal tibia growth plates were stained using TUNEL assay and counterstained with DAPI. Scale bars = 100 μm. b BrdU label retention assays. Sections of the proximal tibia growth plates at different stages were stained for BrdU and Ki-67 and co-stained with DAPI. r resting zone, p proliferative zone. Scale bars = 100 μm. c Experiment design of BrdU label-retention assays. Five doses of BrdU at 50 mg·kg−1 were administered on each day from P12 to P16. Mice were sacrificed at P17, P25, P30, and P43 and the bones were collected for analysis. d Quantification of the number of label-retaining cells (LRCs) per millimeter growth plate in control and cKO growth plates at P30. **P < 0.01. paired t-test. mean ± s.e.m. n = 3 mice for each group
Previous studies have shown that mature or adolescent resting zone of growth plates harbor long-term progenitor cells, which is different from neonatal growth plates.3,4 We found no difference between neonatal cKO and control resting zones as for the shape and number of chondrocytes. However, the resting zone of mature cKO growth plates became significantly thicker than that of the control (Figs. 6b and 3a, b). To test if the chondroprogenitors increased in the resting zone of cKO growth plates, we carried out BrdU label retention assay, which marks putative stem cells (Fig. 6c).4,34 We detected more label-retaining cells (LRCs, putative stem cells) in mature cKO growth plates compared to that of the control (Fig. 6b, c). The resting zones of control growth plates were less than 50 μm in thickness from P17 to P43, while the thickness of cKO growth plates was around 100 μm. At P30, the number of LRCs in cKO growth plates doubled compared to that of control (Fig. 6d). These results demonstrate that the resting zone of mature cKO growth plates becomes dilated, implying a possible role of α-parvin in regulating skeletal stem cell niches.
Single-cell RNA-seq analysis of growth plate chondrocytesTo determine the effects of α-parvin deficiency on the transcriptome of chondrocytes in each distinct zone within the growth plates, we performed single-cell RNA-seq (scRNA-seq) analyses of the control and cKO growth plates. The Uniform Manifold Approximation and Projection (UMAP) dimension reduction and clustering of scRNA-seq data successfully grouped the cells into several distinct zones of the growth plates (i.e., hypertrophic zone, resting zone, proliferative zone, and perichondrium) (Fig. S5A). Next, we analyzed the differentially expressed genes (DEGs) for each cluster. The results showed that gene expression was altered in all three distinct zones of the growth plates (Fig. S6, Table S1). Within the DEGs, some have been reported to regulate chondrogenesis/hypertrophy. Specifically, Tgm2 and Fxyd2, the positive regulators of chondrocyte maturation, increased in the cKO hypertrophic zone (Cluster 0) (Fig. S5B). The marker of hypertrophic chondrocytes, Col10a1, also elevated moderately in the lower cKO proliferative zone (Cluster 1) (Table S1). Ccn2/connective tissue growth factor (CTGF), a multifaceted regulator of skeletal growth, was significantly downregulated (75% and 50% decrease in the lower resting zone and upper proliferative zone) in both the proliferative and resting zones (Clusters 1, 2, 4, 5, 7, and 8) of cKO growth plates (Fig. S5B). Additionally, we noticed an upregulation of osteoarthritis/inflammation-related genes (e.g., Ier3, Klf4, Cytl1) and a downregulation of chondroprotective genes (e.g., A2m, C1qtnf3, Matn1, Matn3)35,36,37,38,39,40 in the cKO growth plates (Fig. S5B). Gene ontology analysis of DEGs from the upper proliferative zone (Cluster 2) suggests significant enrichment in cell-matrix adhesion, cytoskeleton, and ECM (Fig. S7). Indeed, several genes (Ezr, Epyc, Enah, Dcn, Cd44, Actg1, Dstn) of these GO terms were altered significantly (Fig. S5B). However, there was no enrichment in other classical pathways of skeletal growth, such as Fgf, Ihh, and Bmp signaling. These data suggest an altered transcriptome of mutant chondrocytes in multiple zones of the growth plates, providing an explanation of at least some aspects of the observed chondrodysplasia phenotype in the cKO mice.
Comments (0)